Clodronate is encapsulated into homogeneous and stable small unilamellar liposomes of mean diameters between 130 and 150 nm. Encapsulation efficiency is 30% resulting in liposomes containing 18 ±2 mg/ml encapsulated clodronate. Immunohistology (Figure, right) shows the highly effective depletion of tumor associated macrophages using the F 4/80 macrophage marker following treatment of tumor bearing mice with CLODROLIP.
CLODROLIP formulations deplete macrophages very effectively and are of high stability. In contrast to other clodronate-liposome formulations CLODROLIP is a significantly improved formulation that can be stored at -80° C for more than one year without loss of depletion activity. The reagent is optimally suited for repetitive, long-term macrophage depletion experiments.
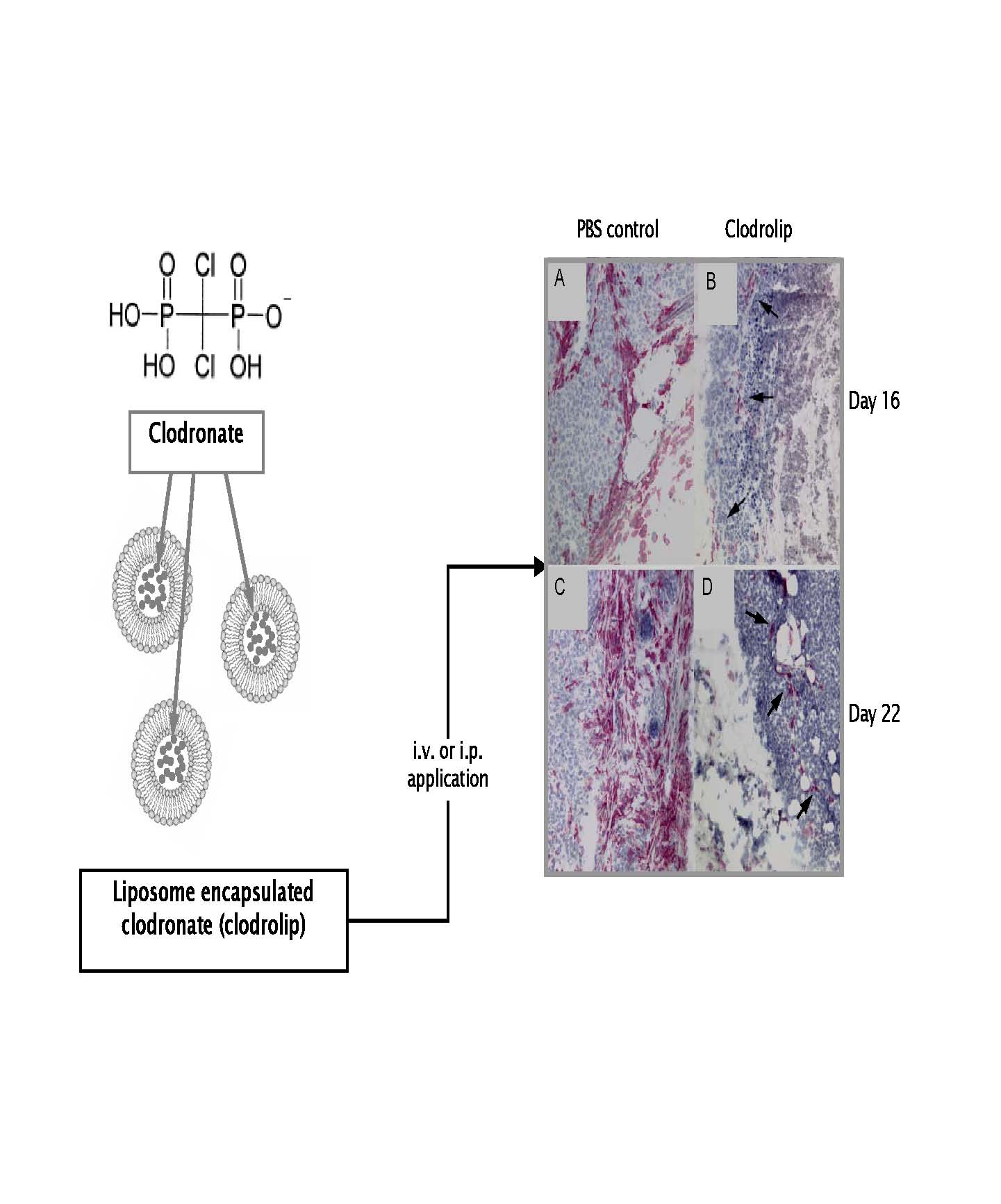
In the figure above CLODROLIP structure and demonstration of macrophage depletion efficiency in A673 rhabdomyosarcoma tumors
(red stained cells are F 4/80+ TAMs) are shown. TAM depletion is >95% (modified from Br. J. Cancer 95: 272, 2006)
(red stained cells are F 4/80+ TAMs) are shown. TAM depletion is >95% (modified from Br. J. Cancer 95: 272, 2006)
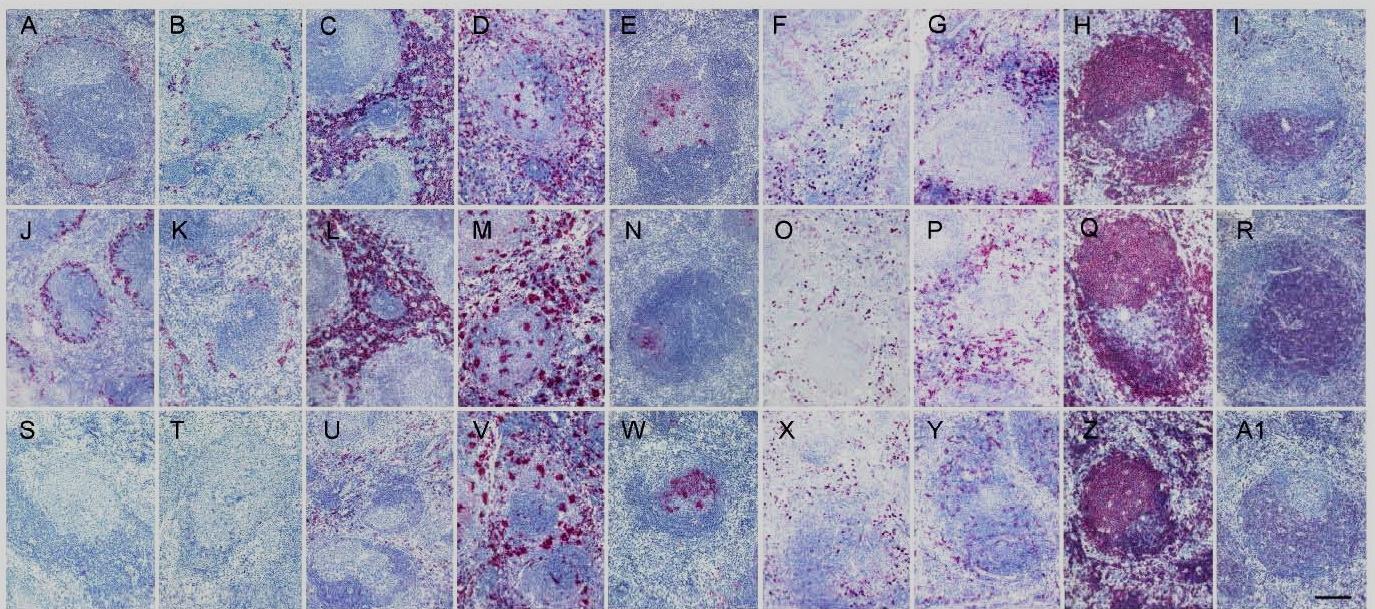
The following figure shows the characteristics of macrophage depletion in the spleen after i.p. treatment with CLODROLIP. Splenic macrophages (red pulp, marginal zone and metallophilic) are fully depleted by CLODROLIP (panels S, T and U), whereas free clodronate has no effect (panels J, K, L and O, P). CD11b+ macrophages and CD11c+ dendritic cells are also depleted but at a lesser extent (panels X and Y). Follicular dendritic cells (FDC), B and T cells are not affected. Interestingly, CD68+ macrophages are not depleted in the spleen of healthy mice (panels D, M and V) or in tumors of tumor bearing mice (see Br. J. Cancer 95: 272, 2006).
Immunohistochemical stains of spleen sections obtained from mice injected with PBS (A to I), with free clodronate (J to R), or with Clodrolip (S to A1):
Here you find PubMed links to recent publications where macrophage depletion was successfully performed using CLODROLIP formulations:
Sequential Activation of Two Pathogen-Sensing Pathways Required for Type I Interferon Expression and Resistance to an Acute DNA Virus Infection. Xu RH, Wong EB, Rubio D, Roscoe F, Ma X, Nair S, Remakus S, Schwendener R, John S, Shlomchik M, Sigal LJ. Immunity 43(6):1148-59, 2015.
Cross-talk between tumors can affect responses to therapy. Devaud C, John LB, Westwood JA, Yong CSM, Beavis PA, Schwendener RA, Darcy PK, Kershaw MH. Oncoimmunology 7: e975572, 2015.
Intravital and whole-organ imaging reveals capture of melanoma-derived antigen by lymph node subcapsular macrophages leading to widespread deposition on follicular dendritic cells. Moalli F, Proulx ST, Schwendener R, Detmar M, Schlapbach C, Stein JV.
Front Immunol. 6:114, 2015. doi: 10.3389/fimmu.2015.00114. eCollection 2015.
ATG16L1 deficiency in macrophages drives clearance of uropathogenic E. coli in an IL-1β-dependent manner. Symington JW, Wang C, Twentyman J, Owusu-Boaitey N, Schwendener R, Nunez G, Schilling JD, Mysorekar IU.
Mucosal Immunol. 8(6):1388-99, 2015.
Inhibition of Cyclooxygenase-2 Prevents Chronic and Recurrent Cystitis. Hannan TJ, Roberts PL, Riehl TE, van der Post S, Binkley JM, Schwartz DJ, Miyoshi H, Mack M, Schwendener RA, Hooton TM, Stappenbeck TS, Hansson GC, Stenson WF, Colonna M, Stapleton AE, Hultgren SJ. eBioMedicine 1, p46-57, 2014.
Heat Shock Protein-Mediated Protection Against Cisplatin-Induced Hair Cell Death. Baker TG, Roy S, Brandon CS, Kramarenko IK, Francis SP, Taleb M, Marshall KM, Schwendener R, Lee FS, Cunningham LL.
J Assoc Res Otolaryngol. 2014 Sep 27.
Macrophage depletion by free bisphosphonates and zoledronate-loaded red blood cells. Sabatino R, Antonelli A, Battistelli S, Schwendener R, Magnani M, Rossi L.PLoS One 9(6):e101260, 2014.
DMXAA causes tumor site-specific vascular disruption in murine non-small cell lung cancer, and like the endogenous non-canonical cyclic dinucleotide STING agonist, 2'3'-cGAMP, induces M2 macrophage repolarization. Downey CM, Aghaei M, Schwendener RA, Jirik FR.
PLoS One. 9(6):e99988, 2014.
Depletion of cutaneous macrophages and dendritic cells promotes growth of Basal Cell Carcinoma in mice.
Koenig S, Nitzki F, Uhmann A, Dittmann K, Theiss-Suennemann J, Herrmann M, Reichardt HM, Schwendener R, Pukrop T, Schulz-Schaeffer W, Hahn H.
PLoS One 9(4):e93555, 2014.
Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation.
Epelman S, Lavine KJ, Beaudin AE, Sojka DK, Carrero JA, Calderon B, Brija T, Gautier EL, Ivanov S, Satpathy AT, Schilling JD, Schwendener R, Sergin I, Razani B, Forsberg EC, Yokoyama WM, Unanue ER, Colonna M, Randolph GJ, Mann DL.
Immunity. 40(1):91-104, 2014.
The depletion of donor macrophages reduces ischaemia-reperfusion injury after mouse lung transplantation. Tsushima Y, Jang JH, Yamada Y, Schwendener R, Suzuki K, Weder W, Jungraithmayr W.
Eur J Cardiothorac Surg. 45(4):703-9, 2014.
Macrophage and T cell produced IL-10 promotes viral chronicity. Richter K, Perriard G, Behrendt R, Schwendener RA, Sexl V, Dunn R, Kamanaka M, Flavell RA, Roers A, Oxenius A.
PLoS Pathog. 9(11):e1003735,2013.
Depletion of resident macrophages does not alter sensory regeneration in the avian cochlea. Warchol ME, Schwendener RA, Hirose K. PLoS One. 7(12): e51574, 2012.
Microparticles released by Listeria monocytogenes-infected macrophages are required for dendritic cell-elicited protective immunity.
Zhang Y, Zhang R, Zhang H, Liu J, Yang Z, Xu P, Cai W, Lu G, Cui M, Schwendener RA, Shi HZ, Xiong H, Huang B.Cell Mol Immunol. 9: 489, 2012.
Macrophages modulate cardiac function in lipotoxic cardiomyopathy. Schilling JD, Machkovech H, Kim AH, Schwendener R, Schaffer JE. Am J Physiol Heart Circ Physiol 303:H1366, 2012.
Inhibition of the Kit ligand/c-Kit axis attenuates metastasis in a mouse model mimicking local breast cancer relapse after radiotherapy. Kuonen F, Laurent J, Secondini C, Lorusso G, Stehle JC, Rausch T, Faes-Van't Hull E, Bieler G, Alghisi GC, Schwendener R, Andrejevic-Blant S, Mirimanoff RO, Ruegg C. Clin Cancer Res. 18(16):4365-4374, 2012.
Radiotherapy promotes tumor-specific effector CD8+ T cells via dendritic cell activation. Gupta A, Probst HC, Vuong V, Landshammer A, Muth S, Yagita H, Schwendener R, Pruschy M, Knuth A, van den Broek M.
J Immunol. 189(2):558-66, 2012.
Dichotomy between RIP1- and RIP3-mediated necroptosis in tumor necrosis factor a-induced shock. Linkermann A, Braesen JH, De Zen F, Weinlich R, Schwendener RA, Green DR, Kunzendorf U, Krautwald S. Mol Med.18(1):577-86, 2012.
Intratumoral macrophages contribute to epithelial-mesenchymal transition in solid tumors.
Bonde AK, Tischler V, Kumar S, Soltermann A, Schwendener R. BMC Cancer 12(1):35, 2012.
Epoxyeicosanoids stimulate multiorgan metastasis and tumor dormancy escape in mice. Panigrahy D, Edin ML, Lee CR, Huang S, Bielenberg DR, Butterfield CE, Barnés CM, Mammoto A, Mammoto T, Luria A, Benny O, Chaponis DM, Dudley AC, Greene ER, Vergilio JA, Pietramaggiori G, Scherer-Pietramaggiori SS, Short SM, Seth M, Lih FB, Tomer KB, Yang J, Schwendener RA, Hammock BD, Falck JR, Manthati VL, Ingber DE, Kaipainen A, D'Amore PA, Kieran MW, Zeldin DC. J Clin Invest. 122(1):178-91, 2012.
S100A7 enhances mammary tumorigenesis through upregulation of inflammatory pathways. Nasser MW, Qamri Z, Deol YS, Ravi J, Powell CA, Trikha P, Schwendener RA, Bai XF, Shilo K, Zou X, Leone G, Wolf R, Yuspa SH, Ganju RK. Cancer Res. 72(3):604-15, 2012.
Inflammation is necessary for long-term but not short-term high-fat diet-induced insulin resistance. Lee YS, Li P, Huh JY, Hwang IJ, Lu M, Kim JI, Ham M, Talukdar S, Chen A, Lu WJ, Bandyopadhyay GK, Schwendener R, Olefsky J, Kim JB. Diabetes 60:2474-2483, 2011.
Albumin coated liposomes: a novel platform for macrophage specific drug delivery. Vuarchey C, Kumar S, Schwendener R. Nanotechnology Development 1:e2, 2011.
Soluble vascular endothelial growth factor receptor-3 suppresses lymphangiogenesis and lymphatic metastasis in bladder cancer. Yang H, Kim C, Kim MJ, Schwendener RA, Alitalo K, Heston W, Kim I, Kim WJ, Koh GY. Mol. Cancer 10(1):36, 2011.
Brain infiltration of leukocytes contributes to the pathophysiology of temporal lobe epilepsy. Zattoni M, Mura ML, Deprez F, Schwendener RA, Engelhardt B, Frei K, Fritschy J-M. J. Neuroscience 31(11):4037-4050, 2011.
Nitric oxide short-circuits interleukin-12-mediated tumor regression. Egilmez NK, Harden JL, Virtuoso LP, Schwendener RA, Kilinc MO. Cancer Immunol Immunother. 60(6):839-45, 2011.
CD11c+ Dendritic cells and B cells contribute to the tumoricidal activity of anti-DR5 antibody therapy in established tumors. Haynes NM, Hawkins ED, Li M, McLaughlin NM, Hämmerling GJ, Schwendener R, Winoto A, Wensky A, Yagita H,Takeda K, Kershaw MH, Darcy PK, Smyth MJ. J Immunol. 185(1):532-41, 2010.
Inadequate clearance of translocated bacterial products in HIV-infected humanized mice.
Hofer U, Schlaepfer E, Baenziger S, Nischang M, Regenass S, Schwendener R, Kempf W, Nadal D, Speck RF. PLoS Pathog. 6(4):e1000867, 2010.
In vivo bioluminescence imaging and histopathopathologic analysis reveal distinct roles for resident and recruited immune effector cells in defense against invasive aspergillosis. Ibrahim-Granet O, Jouvion G, Hohl TM, Droin-Bergere S, Philippart F, Kim OY, Adib-Conquy M, Schwendener R, Cavaillon JM, Brock M. BMC Microbiol. 10(1):105, 2010.
Toll-like receptor triggering and T-cell costimulation induce potent antitumor immunity in mice. Westwood JA, Haynes NM, Sharkey J, McLaughlin N, Pegram HJ, Schwendener RA, Smyth MJ, Darcy PK, Kershaw MH. Clin Cancer Res 15(24):7624, 2009.
Role of CD11b+ macrophages in intraperitoneal lipopolysaccharide-induced aberrant lymphangiogenesis and lymphatic function in the diaphragm. Kim KE, Koh YJ, Jeon BH, Jang C, Han J, Kataru RP, Schwendener RA, Kim JM, Koh GY. Am. J. Pathol. 175(4):1733-1745, 2009.
Myeloid cells contribute to tumor lymphangiogenesis. Zumsteg A, Baeriswyl V, Imaizumi N, Schwendener R, Ruegg C, Christofori G. PLoS One 4(9):e7067, 2009.
Identification of a subpopulation of macrophages in mammary tumor-bearing mice that are neither M1 nor M2 and are less differentiated. Torroella-Kouri M, Silvera R, Rodriguez D, Caso R, Shatry A, Opiela S, Ilkovitch D, Schwendener RA, Iragavarapu-Charyulu V, Cardentey Y, Strbo N, Lopez DM. Cancer Res. 69: 4800-09, 2009.
Critical role of CD11b+ macrophages and VEGF in inflammatory lymphangiogenesis, antigen clearance, and inflammation resolution. Kataru RP, Jung K, Jang C, Yang H, Schwendener RA, Baik JE, Han SH, Alitalo K, Koh GY. Blood 113: 5650-59, 2009.
IL-12 p80-dependent macrophage recruitment primes the host for increased survival following a lethal respiratory viral infection. Gunsten S, Mikols CL, Grayson MH, Schwendener RA, Agapov E, Tidwell RM, Cannon CL, Brody SL, Walter MJ. Immunology 126(4):500-13, 2009.
A pro-inflammatory signature mediates FGF2-induced angiogenesis. Andrés G, Leali D, Mitola S, Coltrini D, Camozzi M, Corsini M, Belleri M, Hirsch E, Schwendener RA, Christofori G, Alcami A, Presta M. J. Cell. Mol. Med. 13(8b): 2083-2108, 2009.
Gene transfer may be preventive but not curative for a lysosomal transport disorder. Hippert C, Dubois G, Morin C, Disson O, Ibanes S, Jacquet C, Schwendener R, Antignac C, Kremer EJ, Kalatzis V. Mol Ther. 16(8):1372-81, 2008.
Deficiency of decay-accelerating factor and complement receptor 1-related gene/protein y on murine platelets leads to complement-dependent clearance by the macrophage phagocytic receptor CRIg. Kim DD, Miwa T, Kimura Y, Schwendener RA, van Lookeren Campagne M, Song WC. Blood 112(4):1109-19, 2008.
Persistent activation of an innate immune response translates respiratory viral infection into chronic lung disease. Kim EY, Battaile JT, Patel AC, You Y, Agapov E, Grayson MH, Benoit LA, Byers DE, Alevy Y, Tucker J, Swanson S, Tidwell R, Tyner JW, Morton JD, Castro M, Polineni D, Patterson GA, Schwendener RA, Allard JD, Peltz G, Holtzman MJ. Nat. Med. 14(6):633-40, 2008.
Morphine induces defects in early response of alveolar macrophages to Streptococcus pneumoniae by modulating TLR9-NF-{kappa}B signaling Wang J, Barke RA, Charboneau R, Schwendener R, Roy S. J. Immunol. 180(5):3594-600, 2008.
Profound but dysfunctional lymphangiogenesis via vascular endothelial growth factor ligands from CD11b+ macrophages in advanced ovarian cancer. Jeon BH, Jang C, Han J, Kataru RP, Piao L, Jung K, Cha HJ, Schwendener RA, Jang KY, Kim KS, Alitalo K, Koh GY. Cancer Res. 68(4):1100-1109, 2008.
Neuroprotective and axon growth promoting effects of intraocular inflammation do not depend on oncomodulin or the presence of large numbers of activated macrophages. Hauk TG, Muller A, Lee J, Schwendener R, Fischer D. Exp. Neurol. 209(2):469-82, 2008.
Interaction between macrophages, TGF-beta1, and the COX-2 pathway during the inflammatory phase of skeletal muscle healing after injury. Shen W, Li Y, Zhu J, Schwendener R, Huard J. J. Cell Physiol. 214(2): 405-412, 2008.
Macrophage depletion diminishes implant-wear-induced inflammatory osteolysis in a mouse model. Ren W, Markel DC, Schwendener R, Ding Y, Wu B, Wooley PH. J. Biomed. Mater. Res. A 85(4): 1043-51, 2008.
Liposomes in biology and medicine. Schwendener RA. Adv. Exp. Med. Biol. 620:117-28, 2007.
PMID: 18217339
Angiogenic role of LYVE-1-positive macrophages in adipose tissue. Cho CH, Koh YJ, Han J, Sung HK, Jong Lee H, Morisada T, Schwendener RA, Brekken RA, Kang G, Oike Y, Choi TS, Suda T, Yoo OJ, Koh GY. Circ. Res. 100(4):e47-57, 2007.
Alveolar macrophages contribute to alveolar barrier dysfunction in ventilator-induced lung injury. Frank JA, Wray CM, McAuley DF, Schwendener R, Matthay MA. Am. J. Physiol. Lung Cell Mol. Physiol. 291(6):L1191-8, 2006.
Clodronate-liposome-mediated depletion of tumour-associated macrophages: a new and highly effective antiangiogenic therapy approach.
Zeisberger SM, Odermatt B, Marty C, Zehnder-Fjallman AH, Ballmer-Hofer K, Schwendener RA. Br. J. Cancer. 95(3):272-81, 2006.
Suppression of alveolar macrophage apoptosis prolongs survival of rats and mice with pneumocystis pneumonia.
Lasbury ME, Durant PJ, Ray CA, Tschang D, Schwendener R, Lee CH. J. Immunol. 176(11):6443-53, 2006.
CCL5-CCR5 interaction provides antiapoptotic signals for macrophage survival during viral
infection. Tyner JW, Uchida O, Kajiwara N, Kim EY, Patel AC, O'sullivan MP, Walter MJ,
Schwendener RA, Cook DN, Danoff TM, Holtzman MJ. Nat. Med. 11:1180-1187, 2005.
Pulmonary aspiration: New therapeutic approaches in the experimental model. Beck-Schimmer
B, Rosenberger DS, Neff SB, Jamnicki M, Suter D, Fuhrer T, Schwendener R, Booy C, Reyes L,
Pasch T, Schimmer RC. Anesthesiology 103(3):556-566, 2005.
Histological analysis of CD11c-DTR/GFP mice after in vivo depletion of dendritic cells.
Probst HC, Tschannen K, Odermatt B, Schwendener R, Zinkernagel RM, Van Den Broek M. Clin.
Exp. Immunol. 141(3):398-404, 2005.
Alveolar macrophages regulate neutrophil recruitment in endotoxin-induced lung injury.
Beck-Schimmer B, Schwendener RA, Pasch T, Reyes L, Booy C, Schimmer RC.Respir. Res.6(1):61, 2005.
Macrophages of the splenic marginal zone are essential for trapping of blood-borne particulate antigen but dispensable for induction of specific T cell responses. Aichele P, Zinke J, Grode L, Schwendener RA, Kaufmann SHE, Seiler P. J. Immunol. 171(3): 1148-1155, 2003.
Decreased alveolar oxygen induces lung inflammation. Madjdpour C, Jewell UR, Kneller S,
Ziegler U, Schwendener RA, Booy C, Klausli L, Pasch T, Schimmer RC, Beck-Schimmer B. Am.J.
Physiol.Lung Cell Mol.Physiol. 284(2):L360-7, 2003.
Roles of macrophages in measles virus infection of genetically modified mice. Roscic-Mrkic
B, Schwendener RA, Odermatt B, Zuniga A, Pavlovic J, Billeter MA, Cattaneo R. J.Virol.
75(7):3343-51, 2001.
Limited mycobacterial infection of the liver as a consequence of its microanatomical structure
causing restriction of mycobacterial growth to professional phagocytes. Seiler P,
Schwendener RA, Bandermann S, Brinkmann V, Grode L, Kaufmann SH, Aichele P.Infect.Immun.69(12):7922-6, 2001.
Crucial role of marginal zone macrophages and marginal zone metallophils in the clearance of
lymphocytic choriomeningitis virus infection. Seiler P, Aichele P, Odermatt B, Hengartner
H, Zinkernagel RM, Schwendener RA. Eur.J.Immunol. 27(10):2626-33, 1997.
Macrophage Links: